
文献解读
代谢物除了作中间体,还施什么“魔法”改变蛋白?
传统观点认为代谢为细胞(生物体)的生存、增殖等生物过程提供能量、生物量合成基础等等,是一种被动适应以满足细胞需求的行为。近些年来,随着代谢组学与其他技术的结合应用,越来越多的研究发现代谢物不仅是生物化学反应的中间体,而且还可以被感知以激活受体和蛋白激酶信号,同时它们还是改变蛋白质定位、活性、相互作用和稳定性的翻译后修饰(PTM)的一种底物。此外,某些代谢酶的改变一方面通过催化作用或非催化作用改变伴侣蛋白的活性、定位或稳定性。上期绘谱君为大家整理了经典调控模式,而本期将从蛋白质翻译后修饰层面为大家整理信号代谢物非经典调控模式(图1)。
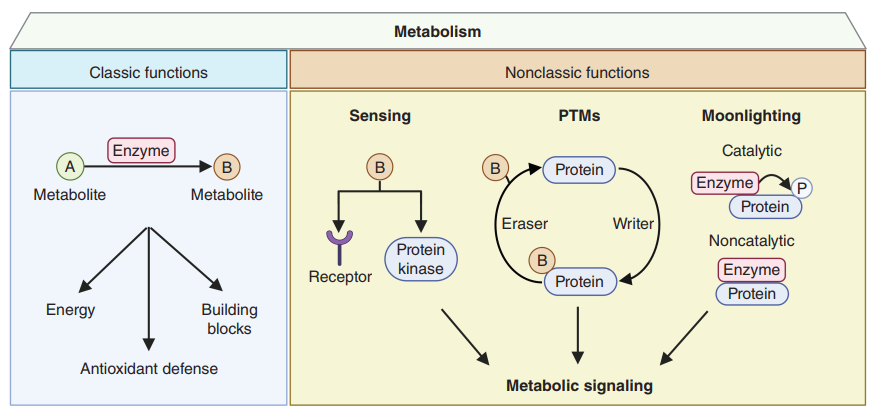
图1. 代谢物调控模式图
代谢物是蛋白质PTMs底物,被称为转移酶的酶促进代谢物与蛋白质共价结合,而擦除器蛋白则撤销这些修饰。此外,蛋白质修饰的程度与可利用代谢物底物的浓度密切相关。代谢物底物浓度和转移酶/擦除剂表达调控PTMs。迄今,已鉴定出600多种PTMs,其中蛋白质磷酸化、甲基化、乙酰化、泛素化、糖基化等是最常见PTMs,此外琥珀酰化、巴豆酰化、2-羟基异丁酰化、乳酸化等是近些年来发现的一系列新型酰化修饰类型。绘谱君按照代谢物种类或者代谢通路为大家总结以下PTMs类型。

图2. 不同PTMs发现时间轴示意图
一、糖酵解-TCA循环代谢物修饰蛋白质
1. 磷酸化(Phosphorylation)
磷酸化是蛋白质激酶催化蛋白质氨基酸残基与三磷酸腺苷(ATP)中磷酸基团之间的连锁反应,可逆性反应则源于蛋白磷酸酶作用去磷酸化。磷酸化通常发生在胞质或细胞核中,被认为是调节蛋白质活性和功能的基本、普遍和基本机制,蛋白质磷酸化是目前研究最深入的PTM之一。根据目标磷酸化的氨基酸残基,蛋白激酶可分为三组:丝氨酸蛋白激酶;苏氨酸蛋白激酶和酪氨酸蛋白激酶。磷酸化位点有丝氨酸、苏氨酸、酪氨酸、半胱氨酸、精氨酸、脯氨酸、天冬氨酸和组氨酸,但最常见的靶位点是丝氨酸(Ser)、苏氨酸(Thr)和酪氨酸(Tyr)。磷酸化可以通过两种机制快速控制蛋白质功能,一种是通过变构来激活酶活性(通常是Ser/Thr,Tyr残基),如糖原合酶激酶3;另一种是通过结合相互作用结构域来激活信号转导(通常是 Tyr 残基)。近30%的人类基因组编码蛋白含有共价结合的磷酸盐,可逆蛋白磷酸化几乎调节与复制、转录、细胞凋亡、免疫反应、环境反应和细胞代谢等细胞过程的各个方面相关。
2. 乙酰化(Acetylation)
蛋白质乙酰化是乙酰转移酶将乙酰辅酶A(Acetyl-CoA)的乙酰基(COCH3)转移到赖氨酸的ε-氨基上。相反,去乙酰化酶催化赖氨酸侧链上乙酰基去除。乙酰化修饰包括一种不可逆型(Nα-乙酰化)和两种可逆型(Nε-乙酰化和O-乙酰化)。这三种类型乙酰化可以发生在不同频率的不同氨基酸上,赖氨酸Nε-乙酰化更常见。目前,人类去乙酰酶有三种类型:Ⅰ类HDAC 由HDAC1、HDAC2、HDAC3和HDAC8组成;Ⅱ类HDAC分为Ⅱa类(HDAC4、HDAC5、HDAC7 和HDAC9)和Ⅱb类(HDAC6和HDAC10);Ⅲ类HDAC是含有SIRT1至SIRT7的烟酰胺腺嘌呤二核苷酸(NAD+)依赖的sirtuins(SIRT)。乙酰化与乙酰辅酶A水平直接相关,线粒体和非线粒体乙酰辅酶A是独立生成。最近研究表明,乙酰辅酶A可以以非酶促方式调节乙酰化。细胞核中组蛋白乙酰化和去乙酰化之间的动态平衡改变了染色质结构并调节基因表达。在过去的十年中,已经鉴定出大量非组蛋白乙酰化,并发现与重要的细胞生物学有关,包括基因表达、DNA 损伤修复、细胞周期控制、细胞命运、蛋白质折叠、蛋白质-蛋白质相互作用、自噬、信号转导和细胞代谢。
3. 琥珀酰化修饰(Succinylation)
琥珀酰化为2004年发现的一种PTM,是指琥珀酰基供体(琥珀酰辅酶A)将琥珀酰基共价结合到底物蛋白质赖氨酸残基的过程。琥珀酰辅酶A是三羧酸(TCA)循环中重要代谢中间体,同时也是琥珀酰化重要供体,越来越多的证据表明,蛋白质琥珀酰化参与转录修饰、免疫反应和细胞代谢,包括TCA循环、尿素循环和脂肪酸代谢。琥珀酰化修饰可发生在细胞核、细胞质和线粒体中。目前研究显示琥珀酰化功能障碍可导致许多疾病,如炎症性疾病、结核病、缺血再灌注样组织损伤和代谢疾病等。
4. 乳酸化修饰(Lactylation)
乳酸化修饰为2019年发现的新型翻译后修饰。酶促作用下,将乳酰辅酶A的乳酰基团结合到蛋白质赖氨酸残基上。非酶促赖氨酸乳酸化是由糖酵解副产物甲基乙二醛衍生而来,产生乳酸谷胱甘肽(LGSH)。乳酸化修饰与多种生理病理过程相关,如细胞分化、M1巨噬细胞极化、线粒体膜结构和功能的稳定性等,同时参与肺纤维化、非酒精性脂肪肝、肾癌、糖尿病、心血管疾病和神经系统失调等疾病的进展。
5. 糖基化(Glycosylation)
糖基化被认为是最丰富和最复杂的PTM,糖基化是在糖基转移酶或糖苷酶作用下将糖复合物(含有聚糖或碳水化合物链)转移至蛋白质氨基酸残基形成糖苷键的可逆酶促过程。糖基化修饰了大约一半的血浆蛋白,而膜和分泌蛋白通常也可被糖基化。糖基化可发生在细胞质、肌膜、内质网和高尔基复合体中。根据连接残基、糖肽键和连接的寡糖,糖基化可分为N-糖基化(与天冬酰胺连接)、O-糖基化(与丝氨酸和苏氨酸连接)、C-糖基化、S-糖基化、糖基磷脂酰肌醇化和磷酸糖基化,O-糖基化和N-糖基化是两种重要的糖基化类型。N-糖基化是最常见的糖基化,其通过β1-糖苷键将N-乙酰氨基葡萄糖(GlcNAc)连接到保守基序Asn-X-Ser/Thr上。O-糖基化将GlcNAc和N-乙酰半乳糖胺(GalNAc)连接到丝氨酸或苏氨酸羟基氧原子上,O-糖基化通常发生在高尔基体翻译后。糖基化在调节细胞过程方面至关重要,包括蛋白质折叠、降解、分泌、分子运输和清除、细胞粘附、细胞间相互作用、信号转导、受体激活和内吞作用。糖基化的失调会影响疾病的发展,包括肿瘤、动脉粥样硬化、糖尿病、肝硬化和阿尔茨海默病。
二、脂肪酸类修饰蛋白质
1. 棕榈酰化(Palmitoylation)
棕榈酸是一种十六碳饱和脂肪酸,可以通过硫酯键连接到半胱氨酸上,它可以增加蛋白质疏水性并促进蛋白质-脂质双分子层相互作用。棕榈酸硫酯反应的不稳定性和可逆性在生理刺激下动态改变蛋白棕榈酰化水平,为调节细胞发育和信号传导提供了关键的可能性。棕榈酰化在神经系统疾病、癌症和代谢紊乱中起着至关重要的作用。
2. 肉豆蔻酰化(Myristoylation)
肉豆蔻酰化是一种不可逆的PTM,由N-肉豆蔻基转移酶(NMT)催化,从肉豆蔻酰辅酶A(Myr-CoA)结合到蛋白N-末端甘氨酸上肉豆蔻酰基化在蛋白质稳定性、蛋白质定位、蛋白质结构成熟、细胞外通讯、免疫反应、细胞代谢和信号转导中至关重要。在癌症、神经系统疾病、病毒和细菌感染以及代谢紊乱中均存在肉豆蔻酰化蛋白失调。
3. 巴豆酰化(Crotonylation)
巴豆酰化于2011年发现的一种修饰种类,是指由巴豆酰基转移酶(HCT)以巴豆酰辅酶A(Cr-CoA)为底物,将巴豆酰基转移到赖氨酸残基上的一种修饰方式(Kcr)。Kcr通常存在于转录活性染色质区域的组蛋白上,与生殖调控密切相关,近年来研究表明,其也可发生在非组蛋白上。赖氨酸巴豆酰化与许多细胞过程有关,包括DNA损伤和修复、干细胞分化、精子发生和炎症。
4. β-羟基丁酰化(β-hydroxybutyryration)
β-羟基丁酰化(Kbhb)是由β-羟基丁酸(β-OHB)介导的一种新型酰化修饰,于2016年被发现。β-OHB为脂质代谢产物,在酰基转移酶p300催化下与赖氨酸结合,而HDAC1和HDAC2可逆地消除Kbhb。kbhb改变的蛋白广泛分布于细胞质、线粒体和细胞核中。之前β-OHB被简单地认为是在饥饿应激下从肝脏向外周组织传递能量的功能性载体。如今β-OHB也是一种重要的信号和表观遗传调控分子,在体内调控生命功能的各个方面。后续研究发现,除了组蛋白外,β-OHB还可以修饰非组蛋白,参与调节癌症、心脏代谢疾病等疾病。
三、氨基酸类修饰蛋白质
1. 甲基化(Methylation)
甲基化是将甲基从活性甲基化合物转移到氨基酸残基的过程,甲基化主要发生在细胞核中,通常修饰核蛋白(如组蛋白),是一种介导DNA转录有效性的表观遗传调控过程。S-腺苷蛋氨酸(SAM)是甲基化反应的底物,它在蛋氨酸循环中产生,而蛋氨酸循环又与叶酸循环和NAD+代谢存在交叉。因此,蛋氨酸和丝氨酸都能增加SAM可用性。蛋白质甲基化可以发生在几个氨基酸残基上,主要修饰赖氨酸和精氨酸。预计人类中存在超过100种赖氨酸甲基转移酶(KMTs),哺乳动物中有9种精氨酸甲基转移酶(PRMTs)。甲基化与各种细胞活动有关,如转录调控、表观沉默、RNA加工和输出、信号转导等。
2. 谷胱甘肽化(Glutathionylation)
S-谷胱甘肽化为谷胱甘肽(GSH)和半胱氨酸巯基(-SH)之间二硫键形成。蛋白质S-谷胱甘肽化是一种可逆的翻译后修饰,可以改变蛋白质结构、活性、亚细胞定位以及与小分子和其他蛋白质的相互作用。谷胱甘肽S-转移酶(GST)可以催化S-谷胱甘肽酰化,或自发发生S-谷胱甘肽酰化。硫氧还蛋白、谷氧还蛋白、硫氧还蛋白硫酸酯酶可以调控谷胱甘肽化逆转。S-谷胱甘肽酰化失衡可导致一系列疾病,如感染、肿瘤、神经退行性疾病、心血管疾病和代谢性疾病。
3. 谷氨酰化(Glutamylation)
谷氨酸可以通过谷氨酰化修饰蛋白质,也称为聚谷氨酰化,是一种可逆的翻译后修饰。该PTM在靶蛋白谷氨酸残基γ-羧基上与酸性谷氨酸结合。谷氨酰化在所有后生动物和原生动物中高度保守,在许多生理和病理过程中发挥关键作用。
四、其他代谢物修饰蛋白质
除了以上介绍的常见代谢物修饰蛋白质以外,共价修饰蛋白质的小分子代谢物还有很多,比如TCA循环中的衣康酸酰化、丙二酰化、戊二酰化、3-羟基丙酰化、亚精胺修饰。短链脂肪酸(SCFA)是肠道中食物消化和膳食纤维发酵的产物,SCFAs可以转化为酰基辅酶A,作为蛋白质赖氨酸酰化供体进行蛋白质修饰如丙酰化、丁酰化、2-羟基异丁酰化、异丁酰化。赖氨酸苯甲酰化(Kbz)是第一个发现的芳香族脂肪酸修饰,主要发生在组蛋白N末端尾部。

小结
代谢物通过PTMs实现了从被动供能到主动调控的转变,动态调节蛋白质功能及细胞命运。代谢物作为PTMs底物的功能失调与多种疾病的发生发展密切相关,如癌症、代谢紊乱和神经退行性疾病。通过总结这些代谢物的非经典调控模式揭示了代谢与表观遗传、免疫及疾病间的深层联系,未来研究需进一步解析PTM修饰与代谢微环境的互作机制,为理解代谢与细胞功能之间的动态互作提供新的视角,为疾病治疗提供新靶点。
参考文献
Wu, X., Xu, M., Geng, M., Chen, S., Little, P. J., Xu, S., & Weng, J. Targeting protein modifications in metabolic diseases: molecular mechanisms and targeted therapies. Signal Transduction and Targeted Therapy, 2023, 8 (1), 220.
请扫描二维码阅读原文

