
文献解读
Cell Reports | 肠道菌群调控环境丰富小鼠的视皮层可塑性

将动物暴露在丰富的环境(enriched environment,EE)中会对动物的大脑结构、功能和可塑性产生显著影响。视觉可塑性是研究大脑神经发育可塑性常用的经典模型,而眼优势可塑性作为研究视皮层可塑性的一个重要模型,其形成原因仍不明确。已有研究发现多种机制有助于EE介导的视皮层可塑性增强,但所有机制都在大脑内部进行研究。来自意大利的科研团队揭示了肠道菌群对EE驱动的视皮层可塑性至关重要,再次验证“肠-脑轴”的重要作用,相关研究成果发表于《Cell Reports》。
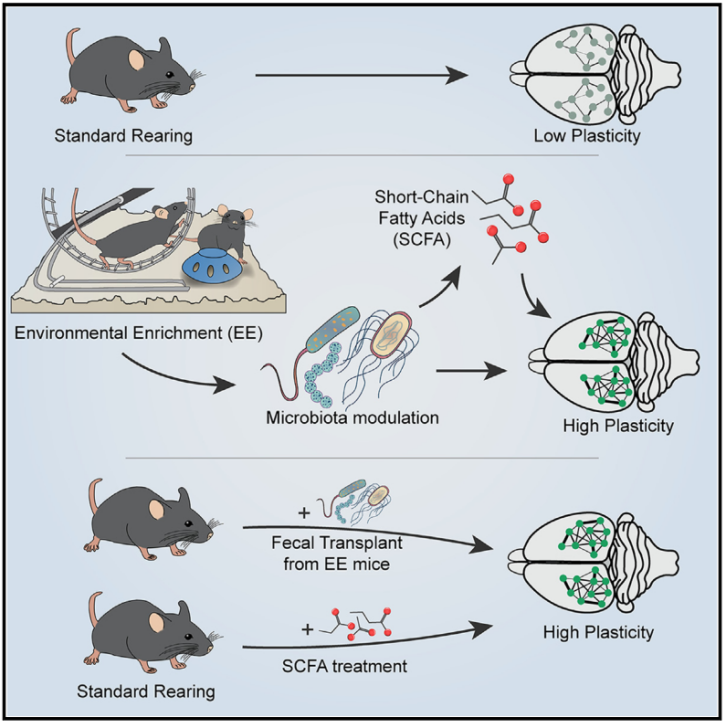
EE影响肠道菌群组成
C57BL/6J小鼠从出生起就在ST(实验室标准笼,仅包含睡眠和筑巢材料)或EE条件下(环境丰富笼,包含跑轮和不同形状的玩具)进行饲养。随后在断奶前,分别收集ST组和EE组出生后第20天(P20),断奶后几天(P25),以及成年期(P90)新鲜粪便进行16S rRNA测序分析。结果显示,与P20小鼠相比,P90小鼠的ST和EE组α多样性均显著增加。而在3个年龄段中,ST组与EE组之间均无显著差异,表明EE饲养不改变菌群α多样性。β多样性的主坐标分析(PCoA)分析表明,ST组小鼠菌群在P20、P25和P90中呈现明显分离趋势。EE组小鼠虽然在P20和P25无显著差异,但在P90和P20之间,P90和P25之间呈现年龄依赖性差异。LEfSe分析结果显示,P20 时,ST小鼠与EE小鼠中某些相对较小的菌群分类群存在显著差异,其中相比于ST组,EE组的Anearoplasmataceae科和Anearoplasmatales目在EE 中显著富集。在P90时, EE小鼠与ST小鼠的菌群组成显著不同。EE组中在P20和P25已经存在的Tenericutes富集继续维持。此外,拟杆菌门中Bacteroidia纲、Bacteroidales目和Rikenellaceae科中的物种在EE中更为丰富。另一方面,属于厚壁菌门的几个分类群在ST中显著富集。与预期一致,PcoA分析显示EE和ST菌群在P90时聚集情况不同,表明 EE和ST小鼠之间肠道菌群组成存在显著差异。总之, EE和ST条件下小鼠之间的菌群组成存在渐进式发展分化。鉴于EE能够在成年期间促进大脑不同区域的不同形式的可塑性,本研究接下来探索了肠道菌群与成年视皮层中EE驱动的可塑性之间的可能联系。
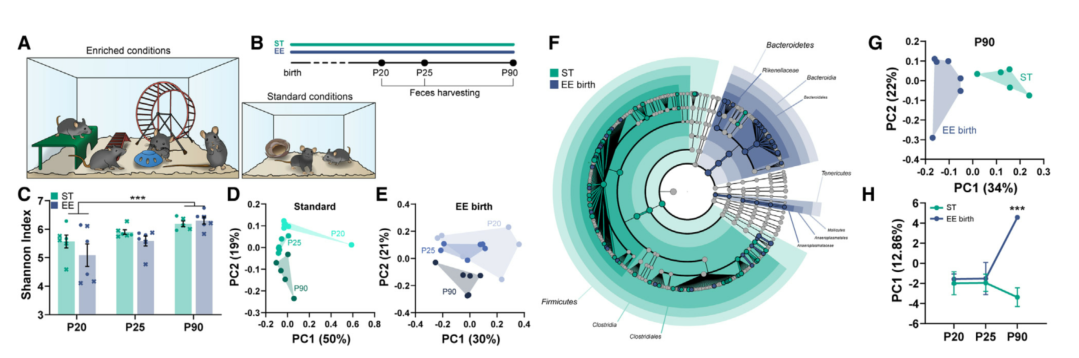
图1. EE调节肠道菌群组成
EE小鼠肠道菌群消耗抑制眼优势可塑性
短时间MD(单眼剥夺)不会诱导成年ST小鼠眼优势可塑性,而在成年EE小鼠中可以诱导强烈的可塑性,表明EE有效地激活了视皮层可塑性。为探究肠道菌群对EE诱导的视皮层可塑性的贡献,在饮用水中使用广谱抗生素(ABX:氨苄青霉素、新霉素、甲硝唑和万古霉素)以耗竭EE小鼠肠道菌群。从出生前1周开始用ABX处理(ABX在母鼠羊水中)并持续到P120。ABX处理后粪便样本的16S rRNA测序结果证明了该处理的有效性。为测试眼优势可塑性,对小鼠进行固有光信号的光学成像(IOS)。在同一受试者MD3天之前和之后(3dMD)分别进行IOS。结果显示ABX处理完全抑制了成年EE小鼠的眼优势可塑性。这些数据表明,完整的肠道菌群对于增强在EE啮齿动物的成年可塑性是必要的。
为排除EE和ABX对发育的影响, P85到P120用EE+ABX处理5周。这种短期方案也足以诱导成年期3dMD引起的眼优势可塑性反应。由于甲硝唑可以穿过血脑屏障,在成年EE小鼠中使用不含甲硝唑(仅氨苄青霉素、万古霉素和新霉素)的ABX。与ABX一样,新型抗生素混合物也能够防止视皮层可塑性。不过,单独使用甲硝唑处理5周并不能阻止成年小鼠EE驱动眼优势可塑性的增强,表明甲硝唑的特定抗菌作用不足以抵消EE可塑性的增强,排除甲硝唑对皮层神经元的直接作用。总之,不受干扰的肠道菌群对于 EE 促进眼优势可塑性是必要的。
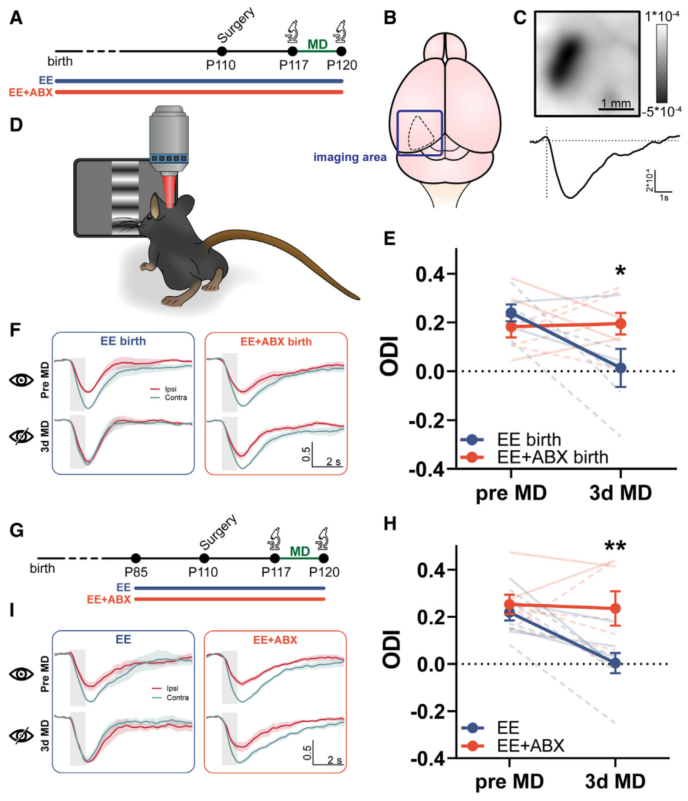
图2. ABX处理抑制EE的可塑性增强作用
ABX处理阻断了EE对树突棘和小胶质细胞形态的影响
树突棘是脑细胞树突中出现的小的膜状突起,有助于向神经元传输电信号。棘突是动态的小隔间,它们的结构重塑被认为是神经元回路正在进行重新布线的标志之一。已知EE会增加皮质树突棘的密度和动态性,那么肠道菌群是否会通过树突棘重塑的机制干扰EE驱动的可塑性。随后在视皮层采集40天内相同树突节段图像。在两个基线成像时间点(间隔1天)后,将小鼠放入EE笼中35天(5周),并摄入含有ABX的水(EE+ABX组)或常规水(EE组),在EE开始后 9、10、19、20、34 和 35 天对两组小鼠相同的树突段进行成像。脊柱密度分析揭示了时间和居住条件之间的显著相互作用。事后检验分析表明,EE小鼠的脊柱密度在所有时间点都显著大于EE + ABX小鼠,但在基线时没有差异。比较各组在不同时间点的脊柱密度发现EE+ABX组的脊柱密度不随时间变化。与EE组基线脊柱密度相比,EE开始后的第一个成像时间点(EE开始后10天)显著增加了25%,而ABX处理组则未增加。为了进一步探究此效应,计算这个关键时间窗口内的脊柱形成和消除率,结果显示,在EE实施的前10天,两组的脊柱消除率无显著差异。然而,与对照组相比,ABX处理组的的脊柱形成率显著降低,解释了ABX处理组的脊柱密度没有增加的原因。同时分析了短期24小时脊柱动力学。结果显示,两组在基线检查时短期形成率相当,但EE处理10天后的短期动力学有显著差异。这些数据表明,ABX处理通过干扰新脊柱形成,而不是破坏现有脊柱稳定性或增加脊柱消除,来阻止EE小鼠树突棘密度的增加。
图3. ABX处理可调节EE对树突棘的影响
视皮层关键期树突棘的眼优势可塑性和活动依赖性调节与小胶质细胞相关。以前研究报告表明,EE诱导不同大脑区域的小胶质突起数量更多、长度更长。那么ABX处理对EE效应的抑制是否也与小胶质细胞形态有关?随后本研究在三组(ST组,EE组,EE+ABX组)的视皮层切片中对 IBA-1(一种特定的小胶质细胞标记物)进行免疫荧光检测。小胶质细胞的定量三维形态分析显示ST和EE小鼠之间存在显著差异,同时,EE小鼠的小胶质细胞复杂性显著增加。而ABX处理完全阻止了EE对小胶质细胞的影响。
为进一步探索其形态学变化,检测了小胶质细胞分支的单一特征,结果显示,与EE组相比,EE+ABX组小鼠形态长度显著缩短,分支点和终点的数量显著减少,而ST小鼠与EE+ABX相似。这些数据表明,ABX处理显著影响EE小鼠的小胶质细胞形态。
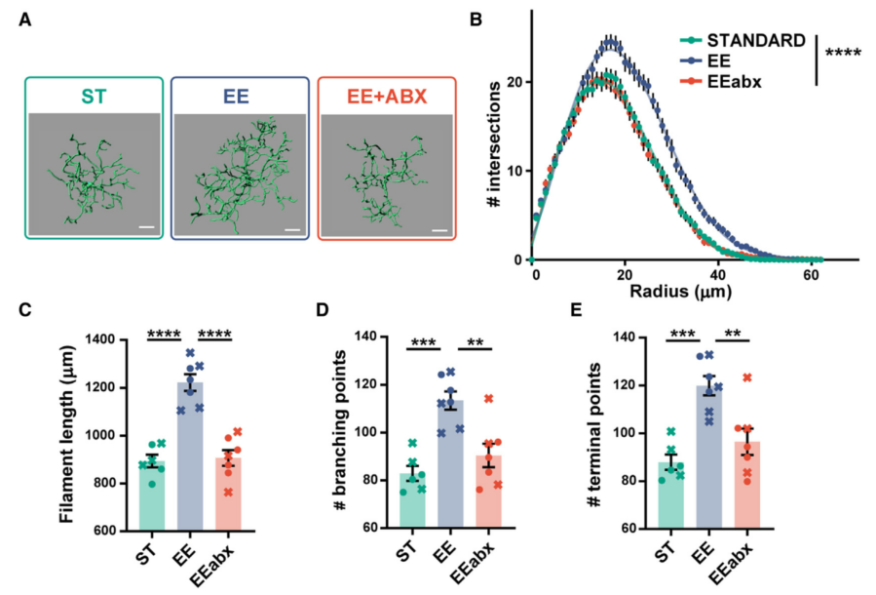
图4. ABX处理改变EE小鼠的小胶质细胞形态
短链脂肪酸处理增强成年小鼠眼优势可塑性并影响小胶质细胞形态
为剖析EE小鼠肠道菌群在成年期促进皮质可塑性的可能机制,随后研究了其共生代谢物。短链脂肪酸(SCFA)是由结肠中菌群发酵膳食纤维和抗性淀粉产生的主要代谢产物,已被证明在神经-免疫-内分泌调节以及肠-脑轴中发挥关键作用。对粪便样本进行SCFA检测,结果显示,与ST小鼠相比,乙酸、丙酸和丁酸浓度在EE组显著增加,而ABX处理完全后这种增加被抑制,暗示SCFA 可能介导EE促进的可塑性效应。将含有乙酸、丙酸、丁酸的SCFA混合物溶解在饮用水中,并给ST成年小鼠服用4周。在处理的最后一周,小鼠在3dMD前后接受IOS成像,结果显示,SCFA促进成年ST小鼠的视皮层可塑性。而且SCFA处理改变了视皮层中小胶质细胞的形态,增加了小胶质细胞过程分枝、细丝长度以及分支点和终点的数量。总之,SCFAs可能是菌群影响视皮层可塑性的候选分子,可能涉及小胶质细胞重塑机制。
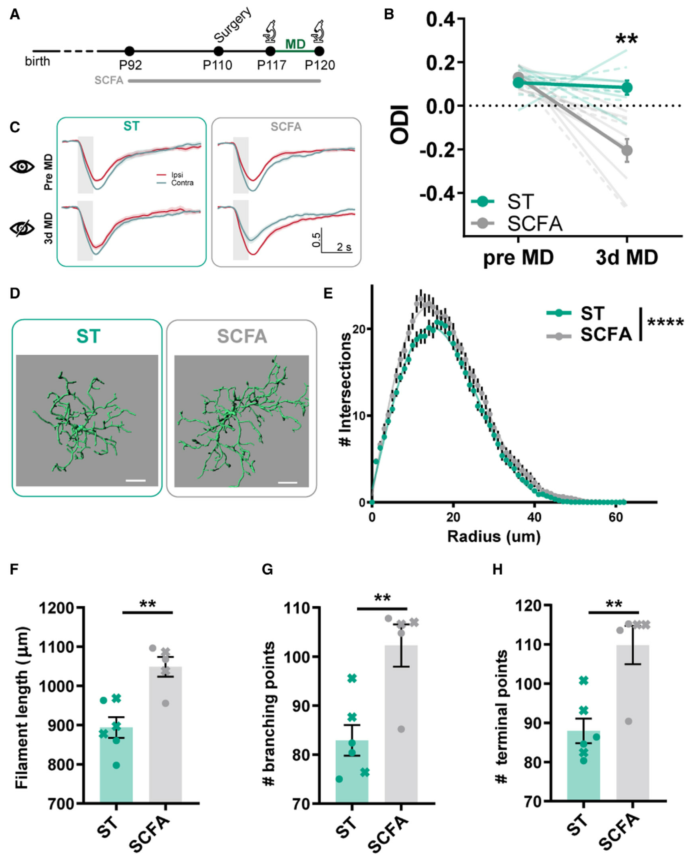
图5.SCFA处理模拟了EE对可塑性和小胶质细胞形态的影响
粪菌移植将EE的可塑性表型转移到ST成年小鼠
收集EE或ST小鼠的新鲜粪便并制成PBS悬浮液,在常规饲养的ST受体小鼠中进行粪菌移植(FT)。4周后,IOS成像研究结果显示, EE供体粪便能够诱导ST受体小鼠的视皮层可塑性,而接受ST供体粪便的ST受体小鼠或PBS对照小鼠在3dMD后未出现任何眼优势转移,与预期一致。
PCoA分析显示,移植EE供体粪便后的ST受体(postFT EE)与EE供体小鼠的菌群呈现重叠,而相对于EE供体和postFT,未移植EE供体粪便的ST受体(preFT EE)呈现显著分离。ANOSIM分析显示,EE供体与 postFT EE组间差异不显著,表明ST受体和EE供体菌群的系统发育组成存在大量重叠。这些数据证明了本研究中FT方案的有效性,并表明成年EE小鼠的可塑性表型可以通过粪便菌群转移到ST小鼠上。
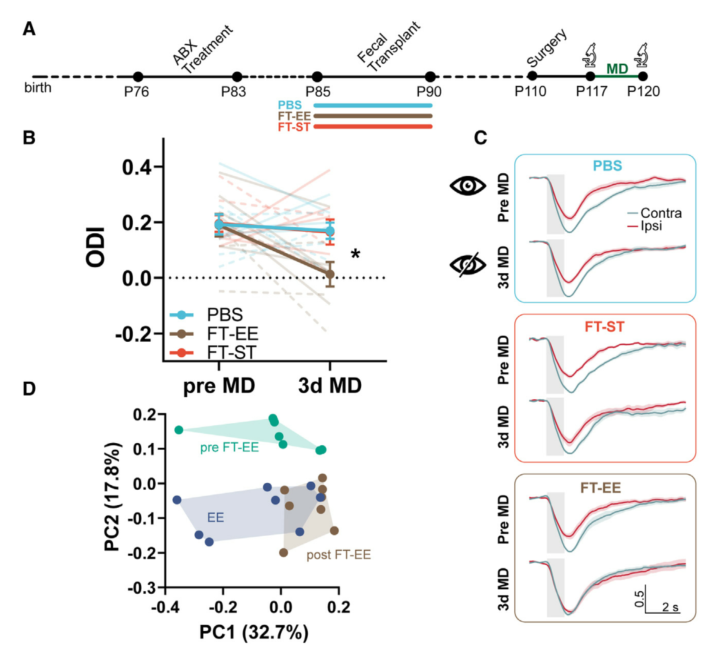
图6.粪便移植ODI
小结
本研究发现EE显著改变小鼠的肠道菌群,且EE小鼠短链脂肪酸(SCFA)水平显著提高。通过对菌群的操纵调控SCFA可干扰EE驱动的视皮层可塑性、树突棘动力学和小胶质细胞重排。而将EE小鼠的粪菌移植到ST受体可激活成年小鼠眼优势可塑性。由此可见,肠道菌群对EE驱动的视皮层可塑性至关重要,再次验证“菌群-肠-脑轴”的重要作用。
参考文献
The gut microbiota of environmentally enriched mice regulates visual cortical plasticity. Cell Reports. 2022.
原文阅读,请扫描下方二维码

绘谱帮你测
短链脂肪酸(SCFAs)是由特定的结肠厌氧菌发酵膳食纤维和抗性淀粉等产生的,是疾病、营养和肠道菌群等之间关键介质。大量报道表明SCFAs与代谢相关疾病(如肥胖、糖尿病)、肠道疾病(如炎症性肠病、结直肠癌)、精神类疾病(阿尔兹海默症、帕金森病、自闭症)有直接或间接的作用。麦特绘谱可绝对定量检测本研究中的短链脂肪酸等代谢物。此外,麦特绘谱拥有业内强大的Q600全定量代谢组、Q300全定量代谢组、Q200宏代谢组、16S rDNA、宏基因组测序等方法,可提供代谢组学一站式整体解决方案,独家的检测技术、全面的数据报告以及专业的售后探讨,助力您的科研探索之路不断创新和突破。详情欢迎咨询麦特绘谱热线400-867-2686,获取详细资料!
往期推荐
1. Nature子刊 | 短链脂肪酸在微生物-肠-脑通讯中的作用(综述)
2. 客户案例 | 中药篇(二):滋补脾阴方药可通过微生物-肠-脑轴改善认知功能
3. SCFAs | 肠道菌群发挥广泛调控作用的“重要武器”–短链脂肪酸

